Gen nhảy là gì?
Các yếu tố vận động, hay “gen nhảy”, lần đầu tiên được xác định bởi Barbara McClintock vào những năm 1940; chúng có thể được gọi chung là transposons. Tại sao transposons lại phổ biến ở tế bào nhân thực đến vậy, và chính xác thì chúng là gì?
Các yếu tố có khả năng vận động (Transposable elements, TEs), còn được gọi là “gen nhảy”, là các trình tự ADN di chuyển từ một vị trí này đến (các) vị trí khác trong hệ gen.
Đang xem: Conjugative transposon là gì, các yếu tố di truyền vận Động
Các yếu tố này lần đầu được xác định trong những thí nghiệm trong giai đoạn 1944 – 1945 bởi nhà di truyền học Barbara McClintock của Phòng thí nghiệm Cold Spring Harbor tại New York. Bà công bố kết quà của mình băm 1951, tuy nhiên các nhà sinh học ban đầu hoài nghi về phát hiện của McClintock.
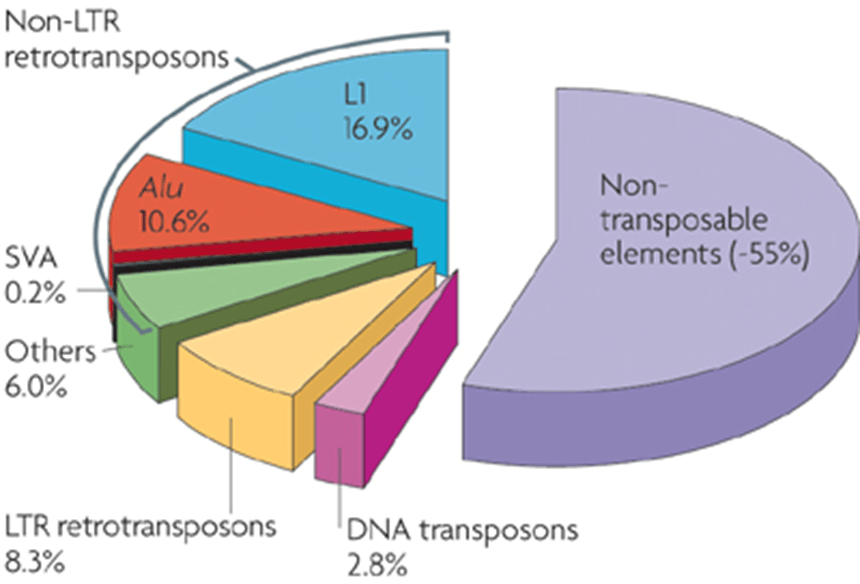
Thế nhưng, qua vài thập kỷ tiếp theo, có vẻ rằng không chỉ có riêng TE “nhảy”, mà chúng còn được phát hiện thấy ở gần như tất cả các sinh vật (cả nhân sơ và nhân thực) và thường có số lượng khá lớn trong hệ gen. Ví dụ, TEs tạo nên gần 50% hệ gen người và tới 90% hệ gen của ngô (SanMiguel, 1996).
Các loại Transposons
Ngày nay, các nhà khoa học biết rằng có nhiều loại TEs khác nhau, cũng như số cách để phân loại chúng.
Một trong số những kiểu phân loại phổ biến đó là giữa các TEs đòi hỏi sự phiên mã ngược (tức là phiên từ ARN thành ADN) để chuyển vị với những TEs không đòi hỏi điều đó. Các yếu tố đầu tiên còn được gọi là retrotransposons hay TEs lớp 1, trong khi nhóm thứ hai còn được biết đến là các DNA transposons hay TEs lớp 2.
Hệ thống Ac/Ds mà McClintock đã phát hiện ra rơi vào nhóm thứ 2. Các nhóm TEs khác nhau được tìm thấy trong hệ gen của các nhân thực khác nhau (hình 1).
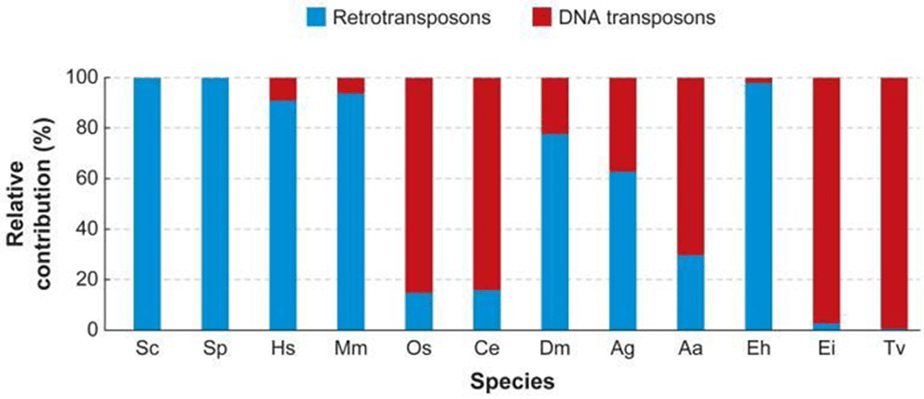
Hình 1: Số lượng tương đối của retrotransposons và DNA transposons trong các hệ gen của loài nhân thực khác nhau. Biểu đồ này cho thấy sự đóng góp của DNA transposons cũng như retrotransposons vào tổng số các loại yếu tố vận động trong hệ gen của các loài. (Sc: Saccharomyces cerevisiae; Sp: Schizosaccharomyces pombe; Hs: Homo sapiens; Mm: Mus musculus; Os: Oryza sativa; Ce: Caenorhabditis elegans; Dm: Drosophila melanogaster; Ag: Anopheles gambiae, malaria mosquito; Aa: Aedes aegypti, yellow fever mosquito; Eh: Entamoeba histolytica; Ei: Entamoeba invadens; Tv: Trichomonas vaginalis.)2007 Annual Reviews Feschotte, C. & Pritham, E. J. DNA transposons and the evolution of eukaryotic genomes. Annual Reviews in Genetics 41, 331–348.
DNA Transposons
Tất cả các TEs lớp 2 hoàn chỉnh hay có thể tự nói là “tự trị” mã hóa protein transposase, thứ mà chúng cần để chèn và cắt (hình 2). Một số trong số các TEs này cũng mã hóa các protein khác.
Chú ý rằng DNA transposon không bao giờ sử dụng RNA trung gian – chúng luôn di chuyển theo cách riêng, cắt và chèn chính mình từ hệ gen sử dụng cơ chế được gọi là “cut and paste”.
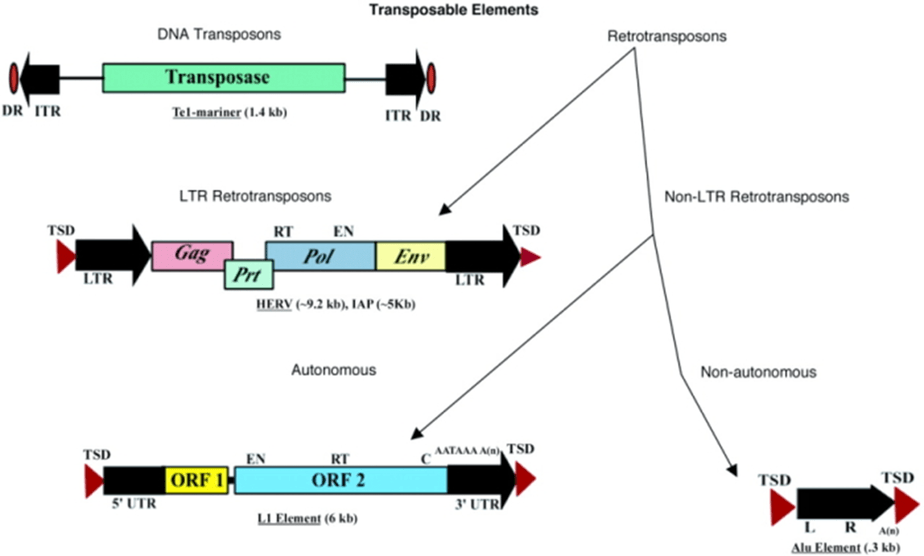
TE lớp 2 được xác định bởi sự có mặt của các trình tự lặp ngược chiều (TIR), khoảng 9 – 40 bp, nằm ở hai đầu (hình 3). Một trong những vai trò của TIR là để được nhận biết bởi transposase.
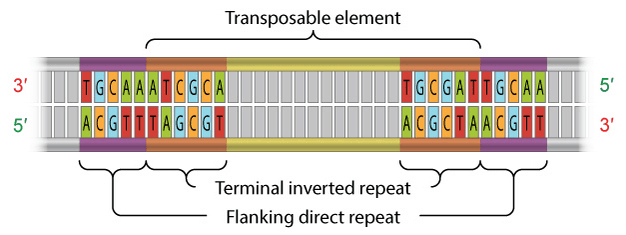
Hình 3: Cấu trúc của một DNA transposon. DNA transposons được bao hai bên bởi TIR. Các trình tự lặp ngược chiều này bổ sung với nhau. 2013 Nature Education Adapted from Pierce, Benjamin. Genetics: A Conceptual Approach, 2nd ed. All rights reserved.
Ngoài ra, cả TE lớp 1 và 2 đều chứa trình tự lặp cùng chiều kẹp hai bên (hình 3). Các trình tự lặp cùng chiều này không phải là một bộ phân thực thụ của TE; thay vào đó, chúng có vai trò trong việc chèn TE vào DNA. Hơn nữa, sau khi một TÉ được cắt, các trình tự lặp này bị bỏ lại như là “những dấu chân”. Đôi khi, những dấu chân này làm thay đổi biểu hiện gen kể cả sau khi yếu tố vận động đã di chuyển đến vị trí khác trong hệ gen.
Dưới 2% hệ gen được tạo nên bởi TE nhóm 2. Điều này có nghĩa là phần lớn trong số các yếu tố vận động thuộc về TEs lớp khác – retrotransposons (Kazazian & Moran, 1998).
Retrotransposons
Không như các yếu tố lớp 2, các yếu tố lớp 1 – tức retrotransposons – di chuyển thông qua hoạt động của RNA trung gian.
Nói cách khác, TEs lớp 1 không mã hóa transposase; thay vào đó, chúng tạo ra các bản phiên mã RNA và sau đó dựa vào enzyme reverse transcriptase để phiên mã ngược trình tự RNA thành DNA, rồi từ DNA mới chèn vào vị trí đích.
Có hai loại chính trong TE lớp 1: LTR retrotransposons, có một đặc trưng là sự có mặt của trình tự lặp dài ở hai đầu (LTRs) ; và non-LTR TEs không có trình tự lặp.
Cả LINE1 (hay L1) và Alu là các họ non-LTR TEs. Yếu tố L1 dài trung bình khoảng 6 kb. Ngược lại, yếu tố Alu thường chỉ chứa mấy trăm nucleotide, vì thế chúng được coi là yếu tố vận động rải rác ngắn, hay short interspersed transposable element, hay SINE.
Alu đặc biệt rất phong phú, vó nguồn gốc từ linh trưởng và trong thời gian tươn đối ngắn nhân lên số lượng thành 1 triệu bản sao trong mỗi tế bào người.
L1 cũng phổ biến trong hệ gen người, mặc dù không có mặt nhiều bản sao như Alu, kích thước lớn của nó có nghĩa rằng yếu tố này tạo nên khoảng 15%–17% trình tự hệ gen (Kazazian & Moran, 1998; Slotkin & Martienssen, 2007).
Ở người, các non-LTR TEs này là nhóm transposon duy nhất còn vận động ; LTR retrotransposons và DNA transposons chỉ còn là những di tích cổ xưa và không còn khả năng di chuyển nữa.
Transposons tự quản và không tự quản
Cả TEs lớp 1 hay lớp 2 đều có thể là tự quản hoặc không.
TEs tự quản có thể tự di chuyển, trong khi các yếu tố không tự quản phải cần sự có mặt của các TEs khác để di chuyển.
Đây là vì các yếu tố không tự quản thiếu các gen cho transposase và reverse transcriptase vốn cần thiết cho quá trình chuyển vị. Ví dụ ở nhân sơ, yếu tố Ac là tự quản vì chúng có thể tự di chuyển, trong khi yếu tố Ds là không tự quản vì phải lệ thuộc vào sự có mặt của Ac.
Gen nhảy làm gì (ngoài việc “nhảy”)?
Thực tế rằng gần một nửa hệ gen người tạo nên bởi TEs, với một tỉ lệ đáng kể chúng là L1 và Alu, thế mới dậy lên một câu hỏi quan trọng : tất cả các gen nhảy này làm gì, ngoài việc nhảy nhót ?
Một transposon làm điều gì phần lớn phụ thuộc vào nơi nó chuyển đến.
Xem thêm: 8 Câu Chúc Ngon Miệng Tiếng Anh Là Gì, Nằm Lòng Những Câu Chúc Ngon Miệng Bằng Tiếng Anh
Nếu nó “hạ cánh” xuống một gen thì có thể gây ra đột biến, như đã được phát hiện khi chèn L1 vào gen mã hóa yếu tố VIII gây bệnh máu không đông (Kazazian et al., 1988).
Tương tự như vậy, một vài năm sau, các nhà nghiên cứu thấy L1 trong gen APC ở các tế bào ung thư dạ dày mà không thấy ở gen APC của những tế bào khỏe mạnh trên cùng một bệnh nhân. Điều này xác nhận rằng L1 chuyển vị trong tế bào thú, và rằng yếu tố này có thể mang một vai trò khởi nguồn của các bệnh (Miki et al., 1992).
Transposons câm
Trái ngược với L1, phần lớn TEs có vẻ câm lặng – nói cách khác, các yếu tố này không tạo ra tác động kiểu hình nào, cũng như không vậ động trong hệ gen. Ít nhất đó cũng là quan điểm đồng tình trong giới khoa học.
Một số TEs câm lặng bị bất hoạt bởi chúng có đột biến tác động đến khả năng di chuyển của chúng ; một số khác hoàn toàn nguyên vẹn và có khả năng vận động nhưng bị giữ bất hoạt do các cơ chế di truyền ngoại gen như là methyl hóa ADN, tái cấu trúc chromatin và miRNAs.
Trong tái cấu trúc DNA chẳng hạn, sự biến đổi hóa học đối với các protein bám chromatin khiến cho chromatin trở nên bị thắt lại ở một số vùng của hệ gen, các gen và cả các TEs trong vùng đó bị bất hoạt vì các enzyme phiên mã đơn giản là không thể tiếp cận promotor gen được.
Một ví dụ khác về transposon câm lặng liên quan đến thực vật, ở chi Arabidopsis.
Các nhà nghiên cứu tập trung đến các thực vật này phát hiện hơn 20 trình tự transposon đột biến khác nhau (trong đó có một loại được phát trên ngô).
Ở các cây kiểu dại, các trình tự này bị methyl hóa, hoặc bị câm lặng. Tuy nhiên, ở các cây mà bị khuyết tận về một hay vài gen tham gia quá trình methyl hóa, các transposon đã được phiên mã.
Hơn nữa, một số kiểu hình đột biến đã được khám phá trong những cây bị khuyết tật quá trình methyl hóa, và các kiểu hình này có liên quan đến sự chèn transposon (Miura et al., 2001).
Dựa vào những nghiên cứu như vậy, các nhà khoa học biết rằng một số TEs bị giữ câm lặng bằng cơ chế ngoại gen ; tuy nhiên, trong những năm gần đây các nhà nghiên cứu bắt đầu tự hỏi liệu chính TEs cũng có vai trò trong quá trình bất hoạt bằng ngoại gen.
Thú vị là, chính Barbara McClintock là người đầu tiên đề suất rằng TEs có thể mang loại vai trò điều hòa này (McClintock, 1951). Cũng phải mất hàng thập kỷ cho các nhà khoa học thu thập bằng chứng để xem xét rằng có vẻ suy luận của McClintock là đúng.
Transposons có thể mã hóa siRNA để điều hòa chính quá trình làm câm lặng chúng
Bởi vì sự chuyển vị của transposon có thể mang tính phá hoại, không có gì ngạc nhiên khi phần lớn trình tự transposon ở hệ gen người là im lặng, vì thế cho phép hệ gen này vẫn tồn tại tương đối ổn định, bất kể tỉ lệ của TEs là thế nào.
Thực tế, các nhà nghiên cứu nghĩ rằng 17% hệ gen người được mã hóa bởi các trình tự liên quan đến L1, chỉ khoảng 100 L1 là vẫn còn hoạt động mà thôi.
Hơn nữa, các nhà khoa học gợi ý rằng kể cả các TEs hoạt động ít ỏi còn sót lại này sự ức chế không thể “nhảy” theo nhiều cách dù cho có vượt qua sự bất hoạt ngoại gen.
Ví dụ, ở tế bào người, small interfering RNAs (siRNAs), còn được gọi là RNAi có thể ngăn cản sự chuyển vị.
RNAi là một cơ chế diễn ra tự nhiên mà sinh vật nhân thực thường dùng để điều hòa biểu hiện gen.
Điều gây thú vị trong tính huống này là siRNA can thiệp vào hoạt tính của L1 là có nguồn gốc từ trình tự 5’UTR của LTR của L1. Cụ thể, 5’UTR của L1 mang một promotor thuận nghĩa cho các gen của L1, cũng như một promotor đối nghĩa mã hóa ra RNA đối nghĩa.
Yang và Kazazian (2006) đã xác định rằng điều này tạo ra các trình tự tương đồng có thể lai với nhau, vì thế hình thành phân tử RNA mạch kép – đáp ứng như là một cơ chất cho RNAi.
Hơn nữa, khi các nhà nghiên cứu ức chế cơ chế làm câm bằng siRNA đó, họ thấy một sự tăng sản phẩm phiên mã L1, gợi ra rằng sự phiên mã yếu tố L1 thực tế bị ức chế bằng siRNA.
Transposon không phải luôn phá hoại
Không phải tất cả sự nhảy chỗ của transopson đều gây ra các tác động có hại.
Thực tế các transposon có thể vận hành quá trình tiến hóa hệ gen bằng cách thúc đẩy sự chuyển vị các trình tự trong hệ gen, sự xáo trộn các exon cũng như sự sửa chữa đứt gãy ADN mạch kép.
Sự chèn và chuyển vị có thể cũng thay đổi các vùng điều hòa gen và các kiểu hình. Thực tế rằng các yếu tố vận động không phải lúc nào cũng có thể cắt một cách hoàn hảo và có thể lấy đi một số trình tự hệ gen làm gây ra hiện tượng được gọi là xáo trộn exon.
Sự xáo trộn exon khiến các exon trước đó không liên quan gì đến nhau nay được xếp kề nhau, thường bằng chuyển đoạn, vì thế có khả năng tạo ra sản phẩm gen mới (Moran et al., 1999).
Xem thêm: ” Tự Hào Là Gì, Làm Thế Nào Để Thoát Khỏi Niềm Kiêu Hãnh, Nghĩa Của Từ Tự Hào
Khả năng của các transposon là tăng đa dạng di truyền, cùng với năng lực của hệ gen trong việc ức chế gần như trọn vẹn hoạt động của TEs, dẫn tới một sự cân bằng, để rồi TEs trở thành một phần quan trọng trong tiến hóa và điều hòa gen trong tất cả các sinh vật có loại trình tự này.
Tham khảo
Bailey, J. A., et al. Molecular evidence for a relationship between LINE-1 elements and X chromosome inactivation: The Lyon repeat hypothesis. Proceedings of the National Academy of Sciences 97, 6634–6639 (2000)Kazazian, H. H. Mobile elements: Drivers of genome evolution. Science 303, 1626–1632 (2004) doi:10.1126/science.1089670Koga, A., et al. Vertebrate DNA transposon as a natural mutator: The medaka fish Tol2 element contributes to genetic variation without recognizable traces. Molecular Biology and Evolution 23, 1414–1419 (2006) doi:10.1093/molbev/msl003McLean, P. McClintock and the Ac/Ds transposable elements of corn, www.ndsu.nodak.edu/instruct/mcclean/plsc431/transelem/trans1.htm (1997)Miki, Y., et al. Disruption of the APC gene by a retrotransposal insertion of L1 sequence in colon cancer. Cancer Research 52, 643–645 (1992)Moran, J. V., et al. Exon shuffling by L1 retrotransposition. Science 283, 1530–1534 (1999)SanMiguel, P., et al. Nested retrotransposons in the intergenic regions of the maize genome. Science 274, 765–768 (1996)
Citation: Pray, L. (2008) Transposons: The jumping genes. Nature Education 1(1):204